
Berberidaceae
| Berberidáceas | ||
|---|---|---|
 | ||
| Taxonomía | ||
| Reino: | Plantae | |
| División: | Magnoliophyta | |
| Clase: | Eudicotyledoneae | |
| Orden: | Ranunculales | |
| Familia: |
Berberidaceae Juss. (1789) nom. cons.[1] | |
| Sinonimia | ||
| ||
Las berberidáceas (Berberidaceae) son una familia de angiospermas perteneciente al orden Ranunculales. Consta de entre 16 y 20 géneros, dependiendo de la inclusión o no de los últimos dos géneros descritos y de otros dos discutidos,[3] y con alrededor de 755 especies aceptadas.[4] Son el grupo hermano de las ranunculáceas, la familia de los ranúnculos, con las que comparten caracteres anatómicos.[5] Se distribuyen, sensu lato, por las regiones templadas y subtropicales. Principalmente en el hemisferio norte y la región andina de Sudamérica.[6]
Muchas de las especies se utilizan ampliamente en jardinería por su valor ornamental.[7] Desde la antigüedad diferentes pueblos las han utilizado en su medicina tradicional para aliviar diversas dolencias, y compuestos derivados de sus principios activos se emplean en la farmacología.[8] En el ámbito culinario, las bayas de algunos miembros del género más numeroso y conocido, Berberis, se consumen frescas, en zumos o mermeladas.[9]
Características
[editar]Esta familia se compone de pequeños árboles y arbustos leñosos perennifolios, además de hierbas perennes rizomatosas o tuberosas, normalmente glabras, raramente con pelos simples unicelulares a uniseriados.
Los tallos pueden ser espinosos o sin espinas,[10] o con espinas nodales como en Berberis. Algunos géneros de la tribu Epimediieae tienen haces vasculares dispersos, con nudos trilacunares o multilacunares, con radios medulares anchos.[6]
Hojas
[editar]El tipo de hojas es muy variado. Pueden ser alternas o basales pero rara vez opuestas; fasciculadas en braquiblastos; perennes o decíduas, de textura herbácea como en el género Epimedium o coriácea; simples, compuestas o recompuestas, a veces con un solo folíolo. Pueden ser enteras, dentadas, espinulosas, lobuladas, palmatífidas o bipartidas, con bases simples a envainantes, o con estípulas menudas a vestigiales, intrapeciolares, caducas, y a veces peltadas. La nervadura también es diversa: pinnatinervia o palmatinervia, a veces modificadas en espinas, como en el género Berberis, y normalmente bifaciales. Con estomas de tipo ranunculáceo solamente en el envés de la hoja.[6]
Inflorescencias
[editar]

Las inflorescencias son terminales, rara vez axilares. Se presentan en racimo, panícula, cima o fascículo, también como flores solitarias o en pareja;[10] bracteadas o sin brácteas, a veces con bracteolas.[nota 1][6]
Las flores son de tamaño pequeñas a medianas; en algunas especies insignificantes y en otras vistosas. Atendiendo a su forma y estructura son bisexuales (perfectas), hipóginas —los elementos de la flor se insertan por debajo del ovario, es decir, con ovario súpero—, con una disposición de esos elementos verticilada o espiralada, básicamente trímeras,[nota 2] o dímeras en el género Epimedium.[11] El receptáculo es corto, no demasiado ahuecado y sin disco ovárico. Normalmente con perianto en número de dos o seis y raramente de nueve verticilos tepaloides —sépalos y pétalos del mismo color y forma—, o con sépalos y estaminodios nectaríferos, o con transición de los estambres a pétalos, que pueden o no ser nectaríferos. En el caso del género Achlys pueden incluso carecer de perianto.[12] En determinados géneros, las flores pueden no tener sépalos, aunque lo normal es tenerlos de 9 o 12 piezas, raramente 50. Normalmente estas piezas se disponen en tres o seis (pocas veces uno y a veces siete) verticilos imbricados de tres piezas cada uno. Según las especies, pueden no tener pétalos, pero lo habitual es que posean 6 o 12 en dos o cuatro verticilos libres de tres piezas cada uno. Frecuentemente los dos o tres verticilos internos tienen nectarios y son interpretados como estaminodios, imbricados, de color amarillo a anaranjado. El androceo tiene tres o seis, raramente 20, estambres trímeros, salvo en Epimedium con estambres dímeros, y se disponen en uno o dos verticilos de tres o seis piezas cada uno. Los estambres son libres y filantéreos, y se disponen de forma opuesta a los tépalos o de forma alterna con los sépalos, según las especies. Las anteras tetraesporangiadas se fijan en la base, con dehiscencia extrorsa o introrsa[nota 3] mediante valvas o hendiduras longitudinales. El gineceo es súpero con un solo carpelo, lo cual es un rasgo característico de la familia, aunque a veces parece que está formado por dos o tres.[13][6] El estilo es corto con estigma apical o subapical, ancho, seco o húmedo, papiloso[nota 4] o no. Suele haber uno u ocho óvulos por carpelo, aunque en ocasiones hay hasta 100, que son anátropos o hemianátropos, bitégmicos, crasinucelados, erectos o péndulos, con placentación de marginal (adaxial) a basal.[6][nota 5]
Fruto y semilla
[editar]Como en las hojas, en el fruto también hay variedad: puede ser carnoso, fundamentalmente en baya, o bien seco y dehiscente de manera irregular en utrículo; o regular en folículo. En el género Caulophyllum, por tener dispersión hidrocórica, no existe fruto al crecer la semilla más rápidamente que la pared del carpelo, que desaparece.[14]
Las semillas pueden ser ariladas o no; a veces con aspecto de drupa azul, como en Caulophyllum, con endospermo oleoso y la capa exterior lignificada —esclerotesta—. El embrión es rudimentario o con radícula larga y cotiledones planos.[6]
- Citología
El número cromosómico es x[nota 6]= 10 en Nandinoideae, x = 8 en Leonticeae, x = 7 en Berberideae y x = 6 en Epimediieae.[15]
Distribución y hábitat
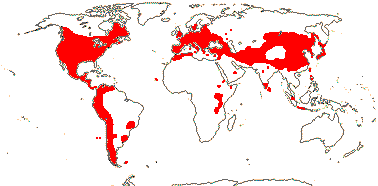
[editar]La mayoría de los géneros (entre 15 y 17) son de distribución original principalmente laurasiana, lo que Good y Tajtadzhián definieron como reino Holártico o reino Boreal (Eurasia y Norteamérica). Berberis, sin embargo, se extiende también por la región andina de Sudamérica y el norte de África, aunque no se conoce aún con exactitud la forma en que se expandieron entre regiones tan alejadas.[16]
Esta expansión intercontinental ha traído de cabeza a botánicos y biogeógrafos desde hace más de un siglo. En 1975, Wolfe expuso su teoría de la «Flora boreotropical» como explicación. En ella sugería que a principios del Terciario el clima del hemisferio norte era cálido uniformemente y las poblaciones se fueron dispersando homogéneamente formando un variado bosque mesofítico durante el Oligoceno y Mioceno, hasta que a finales del Mioceno y durante el Plioceno la temperatura de la Tierra descendió. Como consecuencia, las especies más sensibles y las que habitaban mayores altitudes se extinguieron, o sobrevivieron únicamente en regiones más templadas de Asia, Europa y este de Norteamérica.[17]
Los géneros Bongardia y tres de las especies de Leontice son xerófitos en los ecosistemas áridos y semi-áridos del sureste asiático, el sureste de Europa y el noreste de África. Cinco de las ocho especies de Gymnospermium se encuentran en peligro de extinción y se encuentran amparadas por medidas de protección en Rusia.[18][19]
Epimedium tiene una distribución muy amplia, desde Japón hasta Argelia. Sin embargo, su presencia en algunas regiones de esa vasta extensión asiática y mediterránea es nula. Alrededor del 80% de las especies se concentran en el centro y sureste de China, donde, según ha demostrado un estudio de evolución de los pétalos, se ha mantenido ininterrumpidamente en el tiempo. A pesar de la falta de restos fósiles, se supone que el género se habría extendido ampliamente durante el Paleógeno, o quizás antes de que se produjera el choque de la placa eurasiática con la índica, lo que junto con el posterior levantamiento de la cordillera del Himalaya provocó un cambio en el clima de Asia Central volviéndolo más árido, lo que conllevaría la separación de poblaciones.[20]
Berberis, un género complejo debido a su alta variabilidad morfológica, posiblemente a causa de la hibridación y adaptación al medioambiente, se encuentra representado por 55 especies en India, la mayoría en la región de los Himalayas. Seis de ellas endémicas del estado de Uttarakhand, con B. lambertii en estado de vulnerabilidad, B. affinis y B. osmastonii se consideran raras (Nayar and Sastry 1988) y B. rawatii en peligro de extinción.[21]
El centro de diversidad secundario de Berberis se encuentra en Sudamérica. Chile y Argentina albergan 20 especies endémicas en un área geográfica comprendida entre el desierto de Atacama al norte, el desierto patagónico al este y el océano al sur y oeste. Es una zona compartida por muchas otras especies de leñosas de diferentes géneros, también endémicas. Algunas especies de Berberis se distribuyen por áreas muy restringidas, como B. comberi, que solo se encuentra en algunas localidades de Neuquén y Mendoza (Argentina), B. congestiflora, que habita únicamente el valle central de Chile (entre las regiones de La Araucanía y Los Lagos) o B. corymbosa, endémica del archipiélago Juan Fernández.[22]
Ecología
[editar].jpg)


Las especies leñosas forman parte de setos y espinares, sobre todo en bosques aclarados, en climas continentales de sustratos básicos o calizos.[23]
Estudios fitopatológicos han demostrado que las especies de Berberis son anfitriones alternativos de los hongos causantes del patógeno de la roya. Puccinia graminis (roya negra) y Puccinia striiformis f. sp. tritici (roya rayada o amarilla) son los huéspedes más comunes de estas especies, además de P. brachypodii, P. pygmaea, P. montanensis y P. brachypodii-phoenicoidis, entre otros.[24] Los hongos pasan parte de su ciclo vital en estas plantas sin afectarlas, dispersándose en forma de esporas para colonizar diferentes cultivos de cereales, como el trigo, el centeno, la cebada o la avena.[25] Berberis vulgaris se considera el anfitrión más importante de P. graminis, por lo que normalmente se elimina de las cercanías de los cultivos de trigo para evitar su infección.[26]
Palinología
[editar]El polen se dispersa en mónadas,[27] es tricolpado y tectado-columelado, excepto en Berberis y Ranzania; con 6 o 12 aperturas y estructura parietal amorfa o espiraperturado; con superficie espinosa en Diphylleia, espinulosa en Podophyllum, díadas en Ranzania y tétradas en Sinopodophyllum.[28]
Polinización y dispersión
[editar]La polinización se efectúa de diferentes formas según los géneros. Los estambres de Berberis, Mahonia y Razania son muy irritables y al menor contacto disparan el polen, lo que también facilita su propagación.[14]
La dispersión de las semillas también es variada: las de Caulophyllum se dispersan flotando en el agua (hidrocoria), las de algunas especies de Leontice presentan adaptación a la dispersión como plantas rodantes en los desiertos del sudoeste de Asia, mientras que las semillas de otros géneros tienen una dispersión entomófila, es decir por insectos. Estas semillas están rodeadas por una sustancia aceitosa llamada eleosoma que atrae las hormigas que las arrastran bajo tierra a sus galerías para consumir la capa aceitosa y abandonan la semilla, facilitando de este modo su germinación.[29] Otras, como las del género Vancouveria son dispersadas por la avispa común.[14]
Fitoquímica
[editar]La familia presenta saponinas triterpenoides, numerosos tipos de alcaloides, siendo el más común la berberina, una bencilisoquinolina que tiñe la planta de amarillo. La protopina, un alcaloide aporfínico, está presente en Nandina y la darvasamina, una quinolizidina, en Gymnospermium. Aparecen además flavonoles (miricetina, quercetina, kaempferol) y flavonoides isoprenilados. No son cianogenéticas, excepto Nandina, con compuestos derivados de la tirosina. En Berberis y Leontice se registra la fisiología C3.[nota 7][7]
Usos
[editar]Medicinal
[editar]Numerosas especies de esta familia se utilizan en medicina tradicional, sobre todo en Asia oriental.
En 2015 la comisión farmacéutica de China aprobó el uso de las hojas de Epimedium wushanense como fármaco con el nombre de «Wushan Yinyanghuo». Otra de las especies del mismo género, E. folium, se usa para tratar dolencias óseas y musculares, además de en cosmética.[31]
Las especies de Podophyllum tienen una larga historia en sistemas de medicina tradicional como en las ayurveda, yunani, siddha y entre las tribus nativas norteamericanas, para tratar diversas dolencias. Podophyllum hexandrum, distribuida en la región de los Himalayas, es una especie amenazada debido a la gran demanda en el mercado por su alto contenido en podofilotoxina, de la que se extraen derivados para terapias antitumorales, antivirales, y antifúngicas.[32]
También las especies de Berberis tienen propiedades farmacológicas con diferentes efectos, como antimicrobiano, antiemético, antipirético, antioxidante, antiinflammatorio o sedante.[8] De la más conocida y cultivada, Berberis vulgaris (agracejo), se consumen sus bayas frescas, en zumo, helados o confituras. De otras partes de la planta se hacen infusiones.[9]
Ornamental
[editar]Muchas de las especies de Berberis[33] y Mahonia[34] se usan habitualmente en jardinería como arbustos ornamentales por la vistosidad y colorido de su follaje.[7]
Nandina domestica, la única especie de este género, suele usarse como planta de interior.[35]
Taxonomía
[editar]Posición sistemática
[editar]El sistema de clasificación APG IV de 2016 reconoce a las berberidáceas y las clasifica en el orden Ranunculales en el clado de las eudicotiledóneas.[36] Su estructura floral trímera (en tres partes o múltiplos de tres) y las hojas frecuentemente compuestas son reminiscentes de las lardizabaláceas, con las que han sido normalmente relacionadas, pero de las que difieren por otros caracteres importantes. Basándose en datos moleculares y morfológicos, el Angiosperm Phylogeny Website (APW) considera que forma parte del orden Ranunculales, siendo el grupo hermano de las ranunculáceas[5] (con las que comparte diversos caracteres anatómicos) en un clado moderadamente avanzado del orden.[37]



.JPG)
.jpg)

_N._domestica_-_fl_-_06.jpg)

.jpg)
En los sistemas clásicos se distinguen cuatro grupos de géneros, a los que habitualmente se ha considerado tribus o familias distintas, según el sistema. En la mayoría de los sistemas, el género Nandina es considerado como el más primitivo de la familia y merecedor de una subfamilia propia, Nandinoideae. Los análisis filogenéticos moleculares más recientes que usan el gen cloroplastidial nhdF indican la existencia de tres grandes grupos monofiléticos (que pueden considerarse tribus), bien caracterizados por sus números cromosómicos básicos (x = 6, Epimedieae; x = 7, Berberideae; x = 8, Leonticeae; x = 10, Nandineae), cuyas relaciones no están claras, salvo en el caso de Nandina, que sería el grupo hermano de Caulophyllum (taxones x = 8 que incluyen Caulophyllum, Gymnospermium y Leontice).[38] No queda clara la situación del género Mahonia, que aparece como claramente parafilético y basal del género Berberis[16] y del que se diferencia con dificultad, existiendo además hibridación entre ellos, por lo que ha pasado a considerarse un sinónimo de este.
Subdivisión
[editar]Berberidaceae ha estado sujeta a varias reclasificaciones según diferentes autores. Hutchinson, en 1973, la desgajó en otras dos familias: Nandinaceae —con su único género Nandina—, y Podophyllaceae, en la que incluyó los géneros herbáceos, manteniendo los leñosos —Berberis y Mahonia— en Berberidaceae. Ya en el siglo XXI, el botánico estadounidense Thorne, en sucesivas publicaciones de 2003, 2006 y 2007, reconocía cuatro subfamilias: Nandinoideae, Berberidoideae, Leonticoideae y Podophylloidae. Como resultado de los estudios morfológicos y de ADN, los sistemas de clasificación posteriores, como el APWeb en 2008, volvieron a reunir todos los géneros en Berberidaceae —considerándola monofilética— y la subdividieron en dos subfamilias: Berberidoideae en la que se incluyeron todos los géneros, excepto Nandina, con su propia subfamilia: Nandinoideae. En 2009, el botánico armenio Tajtadzhián segregó otra familia más: Ranzaniaceae. La versión de 2018 de APWeb reconoce tres subfamilias incluyendo Leonticoideae en Podophylloidae.[39]
Subfamilias y tribus
[editar]- La subfamilia Berberidoideae, que comprende dos tribus: Berberideae y Leonticeae, está a su vez dividida en dos subtribus:[40]
- Tribu Berberideae:
- Subtribu Berberidinae con dos géneros: Berberis y Ranzania, junto con los recién nombrados Alloberberis y Moranothamnus, además de los discutidos Mahonia y Holboellia.
- Subtribu Epimediinae con los siguientes géneros: Achlys, Bongardia, Diphylleia, Dysosma, Epimedium, Jeffersonia, Plagiorhegma y Vancouveria.
- Tribu Leonticeae [41] tiene tres géneros: Caulophyllum, Gymnospermium y Leontice.
- Tribu Berberideae:
- La subfamilia Nandinoideae comprende un género con una sola especie aceptada:[42] Nandina domestica.
- La subfamilia Podophylloideae [43] contiene dos géneros: Podophyllum y Sinopodophyllum.
Géneros
[editar]Según diferentes autores y sus publicaciones, se pueden encontrar dentro de la familia entre 15 y casi 40 géneros. Así, Kubitzki y colaboradores incluyeron en 1993 solo 15 géneros en esta familia.[44] Los Jardines de Kew incluyen 18, permutando Aceranthus —sinónimo de Epimedium— con Holboellia.[45][46] WFO —World Flora Online— también incluye 18.[3] Por su parte, el GRIN presenta 22 géneros pertenecientes a la familia, aunque algunos son considerados sinonimias.[47] Mientras que GBIF lista un total de 39 géneros para esta familia, muchos de ellos también sinónimos.[48]
De los 20 géneros no considerados sinónimos de ningún otro, los dos recién descritos —Alloberberis y Moranothamnus— solo aparecen en las publicaciones más modernas. Además, la posición taxonómica de los géneros Mahonia y Holboellia es discutida: el primero puede estar incluido en Berberis y el segundo se clasifica en ocasiones en la familia Lardizabalaceae,[49][50] por lo que tampoco aparecen en todas las clasificaciones.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
+ El género Mahonia está estrechamente relacionado con Berberis y muchos botánicos lo incluyen en él. Sin embargo, una reciente investigación filogenética basada en el ADN lo ha restablecido, aunque con algunas de las especies transferidas a dos géneros recién descritos: Alloberberis (anteriormente Mahonia sección Horridae) y Moranothamnus (anteriormente Mahonia claireae). Las especies de Mahonia y Berberis pueden hibridar, estos híbridos se clasifican en el género × Mahoberberis.[69]
Clave de identificación
[editar]Para identificar los géneros reconocidos se puede seguir la siguiente clave dicotómica.[70] Se trata de una descripción de las estructuras de la planta que diferencian a unos y otros géneros. Para realizar una correcta identificación se ha de leer cada bloque de descripciones (1, 2, 3...) y elegir uno de ellos («a» o «b»), siguiendo hacia el número o género que se indica a continuación.
1a - Si son arbustos leñosos, continuar leyendo en el punto 2.
1b - Si son hierbas perennes, seguir en el punto 3.
2a - Las hojas están bi o tripinnadas. En la flor, el perianto es blanco, tepaloide —el perianto es de tipo tépalo— y sin nectarios. Las anteras de los estambres no se fijan al centro de su filamento, y se abren por una hendidura longitudinal, es decir tienen dehiscencia por hendiduras longitudinales. Se trata del género Nandina Thunb., (1781) que se distribuye de forma natural por Japón, China e India.
2b - Las hojas son simples o pinnadas solo una vez. El perianto es amarillo, con estaminodios internos petaloides —se asemejan a pétalos—, y un par de nectarios. Los estambres son versátiles, es decir, las anteras se fijan al centro del filamento del estambre. Las anteras tienen dehiscencia por valvas —se abren en dos partes como las cáscaras de las pipas de girasol o las placas de los bivalvos—. Estas características pertenecen al género Berberis L., (1753) que es de distribución holártica y suramericana. Se incluye también aquí al antiguo género Mahonia Nutt., (1818).
3a - Si todas las hojas son peltadas, es decir, el pecíolo se inserta en el centro de la hoja, al menos las juveniles se debe seguir en por el punto 4.
3b - Si todas las hojas tienen el pecíolo marginal (lateral), continuar en el punto 7.
4a - Si las anteras son dehiscentes por valvas, se trata del género Diphylleia Michx., (1803) que se distribuye por Norteamérica oriental, China, Japón y la isla de Sajalín.
4b - Si las anteras son dehiscentes por hendiduras longitudinales, hay que continuar por el punto 5.
5a - Las flores son numerosas, rojas y aparecen en inflorescencia cimosa o cerrada. Son las características del género Dysosma Woodson, (1928), planta distribuida por Asia oriental.
5b - Si las flores son solitarias, blancas o rosas entonces ir al punto 6,
6a - Posee rizomas delgados. Tiene habitualmente de 12 a 18 estambres, aunque raramente pueden contarse 9, 10 u 11. Sus flores son alógamas y los frutos son bayas amarillas, rara vez anaranjadas o castañas. El polen aparece en mónadas —granos de uno en uno—, y estos granos son reticulados. Se trata del género Podophyllum (L., 1753), que se distribuye por Norteamérica oriental.
6b - Posee rizomas gruesos. En cada flor autógama solo hay 6 estambres. Los frutos son bayas rojas. El polen se presenta en tétradas —los granos van de cuatro en cuatro—, y poseen espinitas vistos al microscopio. Es el género Sinopodophyllum T.S. Ying, (1979) que se distribuye por Asia oriental.
7a - Las hojas son pinnadas con al menos cinco foliolos. Es la característica diferenciadora del género Bongardia C.A. Mey., (1831), presente en el suroeste de Asia.
7b - Si las hojas son simples o compuestas pero solo en grupos de tres, ir al punto 8.
8a - Si las hojas son simples o con dos o tres foliolos, continuar leyendo en el punto 9.
8b - Si las hojas compuestas están en grupos de tres, se debe ir al punto 12.
9a - Si las hojas conforman el tallo, es decir, realmente no hay un tallo típico, lo que en botánica se denomina hojas caulinares, tienen pétalos con dos nectarios, los estambres versátiles y el fruto es una baya, entonces es el género Ranzania T. Itô, (1888), un endemismo de Japón.
9b - Si tienen hojas basales que nacen directamente en el suelo, sus flores no tienen pétalos o no tienen nectarios, los estambres no son versátiles y el fruto se presenta en folículo o utrículo hay que ir al punto 10.
10a - Las hojas son trifoliadas. Con numerosas flores formando una espiga y con perianto ausente. El fruto se presenta en folículo con una sola semilla o monospermo. Es el género Achlys DC., (1821), que habita en el noroeste de Norteamérica y en Japón.
10b - Si las hojas son simples o bifoliadas, tiene flores solitarias y escaposas, con perianto y su fruto se presenta en utrículo con entre 20 y 50 semillas entonces continuar en el punto 11.
11a - Las hojas son bifoliadas, no tiene tallos aéreos y las flores blancas aparecen de cuatro en cuatro (tetrámeras). Son los caracteres diferenciadores de Jeffersonia (Barton, 1793). Estas plantas se pueden encontrar en Norteamérica oriental.
11b - Las hojas son simples, con tallos aéreos de 8 a 30 cm y flores de color lila trímeras —de tres en tres—. Se trata del género Plagiorhegma Maxim., (1859), que aparece distribuida por Asia oriental.
12a - Si la planta es pubescente, especialmente en los nudos y los bordes de los foliolos, ir al punto 13.
12b - Si es glabra, incluso en los bordes de los foliolos, continuar leyendo en el punto 14.
13a - Las inflorescencias son trímeras, es decir, se observan grupos de tres en tres flores. Característica diferenciadora del género Vancouveria C. Morren & Decne., (1834), que habita el noroeste de Norteamérica.
13b - Las inflorescencias son dímeras —de dos en dos—. Es el género Epimedium L., (1753), cuya distribución es europea y asiática.
14a - Son plantas que tienen rizoma, inflorescencia cimosa, con brácteas subuladas y flores con calículo. El fruto no está desarrollado por lo que las semillas se encuentran expuestas, aunque tienen aspecto de drupa, de color azul. Características del género Caulophyllum Michx., (1803), distribuido por Norteamérica oriental y Asia oriental.
14b - Si son plantas con tubérculo, con inflorescencia en racimo o panícula, con brácteas foliáceas, flores sin calículo y el fruto sí está desarrollado, exponiendo o no las semillas normales, ir al punto 15.
15a - Son plantas de hoja solitaria y estipulada, con semillas expuestas que tienen testa esculturada. El pericarpo del fruto es cartáceo, apicalmente dehiscente, exponiendo parcialmente las semillas. Estas características pertenecen al género Gymnospermium Spach, (1839), que se encuentra en Europa oriental, Asia.
15b - La planta posee de dos a cuatro hojas envainantes, sus semillas tienen testa lisa, el pericarpo del fruto está inflado, es persistente, no exponiendo las semillas. Es el género Leontice L., (1753), que está presente en el suroeste de Asia y de Europa, y noreste de África.
Notas
[editar]- ↑ Brácteas pequeñas que se encuentran sobre el pedicelo.
- ↑ Cuando las piezas anatómicas de un organismo, en este caso la flor, se presentan en número o múltiplo de tres: sépalos, pétalos, estambres, etc, esta se llama «trímera». «Dímera» cuando las partes son dos.
- ↑ Se abren de dentro afuera de la flor, extrorsas; o de fuera hacia dentro, introrsas.
- ↑ Las papilas son pequeñas protuberancias celulares parecidas a pelos.
- ↑ Estos términos se entenderán mejor con una lectura rápida del artículo sobre el óvulo.
- ↑ La "x" expresa el número básico de cromosomas de una especie.
- ↑ C3 es la abreviatura de la vía metabólica de 3 átomos de carbono, un mecanismo de fijación del carbono que utilizan la mayoría de las plantas vasculares que no poseen ningún tipo de adaptación para reducir la fotorrespiración.[30]
Referencias
[editar]- ↑ «Berberidaceae Juss.». Tropicos (en inglés). Consultado el 12 de marzo de 2021.
- ↑ «Berberidaceae». Agricultural Research Service, National Plant Germplasm System. 2020. Germplasm Resources Information Network (GRIN-Taxonomy). National Germplasm Resources Laboratory (en inglés). Beltsville, Maryland. Consultado el 2 de noviembre de 2020.
- ↑ a b «Lista de géneros de Berberidaceae». World Flora Online (en inglés). Consultado el 7 de marzo de 2021.
- ↑ «Berberidaceae». The Plant List (en inglés). Consultado el 31 de octubre de 2020.
- ↑ a b «Ranunculales Juss. ex Bercht. & J.Presl». Botanical Journal of the Linnean Society (en inglés). Consultado el 31 de octubre de 2020.
- ↑ a b c d e f g Watson y Dallwitz, 1992.
- ↑ a b c Michael G. Simpson (2010). «Diversity and Classification of Flowering Plants: Eudicots». Plant Systematics (en inglés) (2da edición).
- ↑ a b «Phytochemistry and Pharmacology of Berberis Species». US National Library of Medicine National Institutes of Health (NCBI) (en inglés). 2014. doi:10.4103/0973-7847.125517. Consultado el 11 de marzo de 2021.
- ↑ a b «Agracejo (Berberis vulgaris)». Naturaleza educativa. Consultado el 11 de marzo de 2021.
- ↑ a b «Berberidaceae Juss.». World Flora Online (en inglés). Consultado el 31 de octubre de 2020.
- ↑ «Morfología de plantas vasculares - Disposición de las piezas florales». Facultad de Ciencias Agrarias. Consultado el 5 de marzo de 2021.
- ↑ Kubitzki, Rohwer, Bittrich, 1993, «Froral structure», p. 147.
- ↑ Simpson, 2010.
- ↑ a b c Kubitzki, Rohwer, Bittrich, 1993, «Fruit and seed», p. 148.
- ↑ Kubitzki, Rohwer, Bittrich, 1993, «Karyology», p. 147.
- ↑ a b Young Dong Kim, Sung-Hee Kim, Leslie Landrum (julio de 2004). «Taxonomic and phytogeographic implications from ITS phylogeny in Berberis (Berberidaceae)». Journal of Plant Research volumen = 117 (en inglés). pp. 175-182. doi:10.1007/s10265-004-0145-7. Consultado el 5 de marzo de 2021.
- ↑ «Phylogenetic and Biogeographic Diversification of Berberidaceae in the Northern Hemisphere». Systematic Botany (en inglés) (32). 2007. pp. 731-742. doi:10.1043/06-16.1. Consultado el 23 de marzo de 2021.
- ↑ Kubitzki, Rohwer, Bittrich, 1993, «Distribution and Habitats», p. 147-9.
- ↑ «Mapa de distribución». APWeb. Consultado el 6 de marzo de 2021.
- ↑ «Petal evolution and distribution patterns of Epimedium L. (Berberidaceae)». Europe PMC (en inglés). Consultado el 23 de marzo de 2021.
- ↑ Umeshkumar L. Tiwari, Bhupendra Singh Adhikari, Gopal Singh Rawat. «A checklist of Berberidaceae in Uttarakhand, Western Himalaya, India». CheckList - The journal of biodiversity data (en inglés). Consultado el 23 de marzo de 2021.
- ↑ Landrum, Leslie (1999). «Revision of Berberis (Berberidaceae) in Chile and Adjacent Southern Argentina». Annals of the Missouri Botanical Garden (en inglés) 86 (4): 793-834. Consultado el 23 de marzo de 2021.
- ↑ Castroviejo, S. et al., ed. (1986). «XXXVII. Berberidaceae». Flora ibérica: plantas vasculares de la Península Ibérica e Islas Baleares. VOL. I. Lycopodiaceae - Papaveraceae. Madrid. p. 403-405. ISBN 84-00-06221-3. Consultado el 27 de marzo de 2021.
- ↑ Anna Berlin, Annemarie F Justesen, Julia Kyaschenko, Jonathan Yuen (2013). «Rust Fungi Forming Aecia on Berberis spp. in Sweden». Plant disease: 1281-1287. doi:10.1094/PDIS-10-12-0989-RE. Consultado el 17 de marzo de 2021.
- ↑ Yue Jin; Les J. Szabo; Martin Carson (2010). «Century-Old Mystery of Puccinia striiformis Life History Solved with the Identification of Berberis as an Alternate Host». PHYTOPATHOLOGY 100 (5). doi:10.1094/PHYTO-100-5-0432.
- ↑ Singh, Ravi P.; Hodson, David; Huerta-Espino, Julio; Jin, Yue; Njau, Peter; Wanyera, Ruth; Herrera-Foessel, Sybil; Ward, Richard W. (2008). «Will Stem Rust Destroy The World's Wheat Crop?». Advances in Agronomy 98. pp. 272-309. ISBN 9780123743558. doi:10.1016/S0065-2113(08)00205-8. Archivado desde el original el 8 de noviembre de 2020. Consultado el 18 de marzo de 2021.
- ↑ «Mónada». Plantas y Hongos. Consultado el 2 de noviembre de 2020.
- ↑ Kubitzki, Rohwer, Bittrich, 1993, «Pollen morphology», p. 147.
- ↑ Kubitzki, Rohwer, Bittrich, 1993, «Fruit and Seed», p. 148.
- ↑
- ↑ Ting Liua; Xiang Liub; Yanjiao Luob; Yu Yaob; Chunmei Wenb; Qianru Yangb; Guoan Shenband; Baolin Guo (2020). «The complete chloroplast genome of Epimedium wushanense T. S. Ying. (Berberidaceae), a traditional Chinese medicinal herb». Mitochondrial DNA Part B - Resources (en inglés) 5 (1): 817-818. doi:10.1080/23802359.2020.1715877.
- ↑ Om Prakash Chaurasia, Basant Ballabh, Amol Tayade, Raj Kumar (2012). «Podophyllum L.: An endergered and anticancerous medicinal plant-An overview». Indian Journal of Traditional Knowledge 11 (2): 234-241. Consultado el 11 de marzo de 2021.
- ↑ «Barberry». Britannica (en inglés). Consultado el 11 de marzo de 2021.
- ↑ «Oregon grape». Britannica (en inglés). Consultado el 11 de marzo de 2021.
- ↑ «Berberidaceae». Britannica (en inglés). Consultado el 11 de marzo de 2021.
- ↑ The Angiosperm Phylogeny Group (1 de mayo de 2016). «An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV». Botanical Journal of the Linnean Society (en inglés) 181 (1): 1-20. ISSN 1095-8339. doi:10.1111/boj.12385.
- ↑ «An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV». Botanical Journal of the Linnean Society (en inglés). Consultado el 31 de octubre de 2020.
- ↑ Kim Y.-D., Kim S.-H., Kim C.-H., y Jansen R.K. (2004). «Phylogeny of Berberidaceae based on sequences of the chloroplast gene ndhF». Biochemical Systematics and Ecology 32: 291-301.
- ↑ Singh, Gurcharan (2019). CRC Press, ed. Plant Systematics: An Integrated Approach, Fourth Edition (en inglés). p. 370. ISBN 9781000576771.
- ↑ «Berberidoideae». GRIN Global (en inglés). Consultado el 21 de marzo de 2021.
- ↑ «Leonticeae». GRIN Global (en inglés). Consultado el 21 de marzo de 2021.
- ↑ «Nandinoideae». GRIN Global (en inglés). Consultado el 21 de marzo de 2021.
- ↑ «Ranunculales». APW (en inglés). Consultado el 21 de marzo de 2021.
- ↑ Kubitzki, Rohwer, Bittrich, 1993, «Key to the genera», p. 148-9.
- ↑ «List of genera in family Berberidaceae». Vascular Plant Families and Genera. Consultado el 31 de marzo de 2021}.
- ↑ «The Plant List. Gen. Aceranthus». Consultado el 30 de marzo de 2021.
- ↑ «Genera and generic subdivisions of Berberidaceae Juss., nom. cons.». U.S. National Plant Germplasm System.
- ↑ «Berberidaceae Juss., 1789». 2019. The International Plant Names Index Collaborators. International Plant Names Index. Consultado el 30 de marzo de 2021.
- ↑ Christopher Brickell (2019). «RHS Encyclopedia Of Plants and Flowers». DK.
- ↑ «The Plant List, ver. 1». Consultado el 30 de marzo de 2021.
- ↑ «Berberis». eFloras (en inglés). Consultado el 11 de marzo de 2021.
- ↑ «Mahonia». eFloras (en inglés). Consultado el 11 de marzo de 2021.
- ↑ «Ranzania». The Plant List (en inglés). Consultado el 11 de marzo de 2021.
- ↑ «Holboellia». eFloras (en inglés). Consultado el 11 de marzo de 2021.
- ↑ «Nandina». eFloras (en inglés). Consultado el 11 de marzo de 2021.
- ↑ «Caulophyllum». eFloras (en inglés). Consultado el 11 de marzo de 2021.
- ↑ «Gymnospermium». eFloras (en inglés). Consultado el 11 de marzo de 2021.
- ↑ «Bongardia». eFloras (en inglés). Consultado el 11 de marzo de 2021.
- ↑ «Achlys». eFloras (en inglés). Consultado el 11 de marzo de 2021.
- ↑ «Diphylleia». eFloras (en inglés). Consultado el 11 de marzo de 2021.
- ↑ «Dysosma». eFloras (en inglés). Consultado el 11 de marzo de 2021.
- ↑ «Podophyllum». eFloras (en inglés). Consultado el 11 de marzo de 2021.
- ↑ «Sinopodophyllum». eFloras (en inglés). Consultado el 11 de marzo de 2021.
- ↑ «Epimedium». eFloras (en inglés). Consultado el 11 de marzo de 2021.
- ↑ «Vancouveria». eFloras (en inglés). Consultado el 11 de marzo de 2021.
- ↑ «Jeffersonia». eFloras (en inglés). Consultado el 11 de marzo de 2021.
- ↑ «Plagiorhegma». eFloras (en inglés). Consultado el 11 de marzo de 2021.
- ↑ «Leontice». eFloras (en inglés). Consultado el 11 de marzo de 2021.
- ↑ Chih-Chieh Yu, Kuo-Fang Chung. «Why Mahonia? Molecular recircumscription of Berberis s.l., with the description of two new genera, Alloberberis and Moranothamnus». Wiley Online Library (en inglés). Consultado el 11 de marzo de 2021.
- ↑ Theodor C. H. Cole, Julien B. Bachelier (junio de 2019). «Berberidaceae Phylogeny Poster». Project: Ranunculales Phylogeny Posters (en inglés). Consultado el 7 de marzo de 2021.
{kind=link}
Bibliografía
[editar]- Kubitzki, Rohwer, Bittrich, Klaus, Jens G. , Volker (1993). Springer Science & Business Media, ed. Flowering Plants · Dicotyledons: Magnoliid, Hamamelid and Caryophyllid Families (en inglés). pp. 147- 150. ISBN 9783540555094.
- Loconte, H. (1993). «Berberidaceae.». Kubitzki, K., Rohwer, J.G. & Bittrich, V. (Editores). The Families and Genera of Vascular Plants. II. Flowering Plants - Dicotyledons. Springer-Verlag: Berlín. ISBN 3-540-55509-9.
- Simpson, Michael G. (2010). «Diversity and Classification of Flowering Plants: Eudicots - Berberidaceae in Plant Systematic». ScienceDirect (en inglés) (2ª edición). Consultado el 31 de octubre de 2020.
- Watson, Dallwitz, L., M.J. (1992 en adelante). «The families of flowering plants: descriptions, illustrations, identification, and information retrieval. Version: 19th October 2020.». Consultado el 31 de octubre de 2020.
Enlaces externos
[editar]| Control de autoridades |
|
|---|
 Datos: Q155949
Datos: Q155949 Multimedia: Berberidaceae / Q155949
Multimedia: Berberidaceae / Q155949 Especies: Berberidaceae
Especies: Berberidaceae