
Saccoglossus kowalevskii
| Saccoglossus kowalevskii | ||
|---|---|---|
 | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Hemichordata | |
| Clase: | Enteropneusta | |
| Familia: | Harrimaniidae | |
| Género: | Saccoglossus | |
| Especie: |
Saccoglossus kowalevskii Agassiz, 1873 | |
Saccoglossus kowalevskii es una especie de «gusano bellota» de la familia Harrimaniidae, de la clase Enteropneusta y el filo Hemichordata.
Es un animal marino, solitario de vida libre y parecido a los gusanos, con un plan corporal simple y con simetría bilateral, que no pasa por una etapa de larva.
Se puede encontrar en hábitats costeros (zona intermareal) de barro y arena. Cava tubos en el sustrato, expulsando montones cónicos de piezas fundidas en forma de espiral. Se caracteriza especialmente por los anillos concéntricos de fibras musculares en la trompa (proboscide).[1]
Embriología[editar]
La embriogénesis de S. kowalevskii corresponde a la de un organismo con un plan corporal adulto que no atraviesa por la etapa de larva (Desarrollo directo). [2]
Etapas iniciales[editar]
- Blástula

Microscopía electrónica de barrido.
El huevo fertilizado de S. kowalevskii se desarrolla en una etapa temprana de blástula a la edad de 8 horas después de la fertilización, cuando mide aproximadamente 400 micrómetros (μm) de altura y 430 μm de ancho.
- Etapa de gástrula
Después de 24 horas pos-fertilización, la gástrula temprana tiene forma de copa y mide 300 μm de altura y 340 μm de ancho.

A una edad entre 30-36 horas, la mayoría de los embriones desarrollan una forma ovoide alargada de gástrula.
Uno de los caracteres llamativos es el telotroque que está constituido por células multiciliadas que rodean a los animales aproximadamente 50 μm por delante del blastoporo.[3]
- Primera etapa de surco
Los embriones tienen una forma alargada y miden 450 μm de longitud, con el extremo anterior más redondeado y un extremo posterior más aplanado. Este surco (telotroque) que rodea al embrión separa el campo perianal posterior de la parte anterior más grande.
- Etapa temprana de torsión
La abertura bucal ventromediana se ha desarrollado y conecta el endodermo anterior con el entorno exterior.
- Etapa tardía de torsión
Los embriones a las 96 horas pos-fecundación miden alrededor de 470 μm de longitud total y el cuerpo se puede dividir en tres regiones: la región de la probóscide anterior, la región del cuello medio y la región del tronco retorcido.
- Etapa 1 de hendidura branquial
Los embriones de esta etapa miden aproximadamente 510 μm de largo. La epidermis dorsal de la región del collar muestra una depresión longitudinal, el surco neural donde procede la neurulación del cordón del collar.
Etapa juvenil[editar]
- Etapa 2 de hendiduras branquiales

Arriba: distribución interna a través de epidermis transparente.
Abajo: Sistema nervioso aislado (en amarillo).
Estos juveniles se arrastran libremente sobre el sustrato y su apariencia general se asemeja a la de los adultos en muchos aspectos, excepto por el reducido número de poros branquiales, el desarrollo de las gónadas y la presencia de una cola postanal.
El pequeño gusano bellota mide unas 800 μm de largo.[3]
Los juveniles de 14 días después de fertilización presentan 2 a 3 hendiduras branquiales.
S. kowalevskii es un organismo con un plan corporal adulto que no atraviesa por una etapa de larva (Desarrollo directo). El estudio del desarrollo directo de este gusano marino comparada con la de los vertebrados, identificó regiones homólogas de sus planes corporales adultos y también la expresión de un conjunto de factores de transcripción con funciones conservadas en el patrón Antero-posterior.[2]

Proteína bmp2/4 marcada en violeta, ubicada en el cordón dorsal. Inmunohistoquímica.
El S. kowalevskii juvenil se alimenta con su probóscide muscular utilizada también para excavar, un collar y un tronco largo dividido en regiones funcionales discretas a lo largo del eje anteroposterior.
Este gusano vive en los sedimentos en una madriguera-túnel en forma de U con agujeros separados de alimentación y de defecación con espirales cónicas de material fecal.
Los individuos se alimentan en los depósitos bentónicos usando una proboscide muscular y ciliada para sondear la superficie del sedimento cerca de la abertura de su túnel.
[4]
Los gusanos bellota filtran el agua marina a través de unas aberturas situadas entre su sistema digestivo y su esófago. Estas aberturas están relacionadas evolutivamente con las agallas de los peces y representan una innovación crítica en la historia evolutiva que no es compartida por otros animales como las lombrices o los artrópodos.[5]
Anatomía[editar]

El Saccoglossus kowalevskii crece hasta aproximadamente 20 centímetros (cm) de largo.
Muestra una polarización dorsal-ventral en la ubicación de sus órganos, como las hendiduras branquiales, las gónadas y el complejo corazón/riñón.
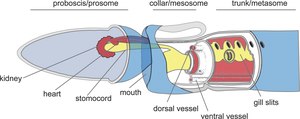
Tiene una red nerviosa y tractos de axones, pero no tiene sistema nervioso central, no tiene una masa de ganglios neurales similar al cerebro y, a diferencia de los cordados y artrópodos, sus células nerviosas y células epidérmicas están finamente entremezcladas.[6]
Sistema Nervioso[editar]
Saccoglossus kowalevskii como hemicordado se diferencia en que carece de sistema nervioso central en forma de ganglios, porque se encuentra dispuesto en forma de plexos nerviosos en el ectodermo.[7] Esto sugiere un «cerebro» bilateral complejo debajo de la piel, con conexiones específicas, formadas por gran cantidad de tipos de neuronas.[8]
En el embrión de S. kowalevskii durante su desarrollo se encuentra un eje molecular que precede al eje dorsoventral de las especializaciones anatómicas y fisiológicas. Expresa el gen Chordin en la línea media ventral y el gen bmp2/4 en la línea media dorsal.

El sistema nervioso de S. Kowalevskii tiene una red nerviosa y tractos de axones, y, sus células nerviosas y células epidérmicas están finamente entremezcladas.[6] Esto sugiere un «cerebro» bilateral complejo debajo de la piel, con conexiones específicas, formadas por una gran cantidad de tipos de neuronas con funciones específicas.[8]
Este sistema consta de un plexo nervioso concentrado en la parte anterior, junto con cordones nerviosos tanto en el lado dorsal como en el ventral.[9]


También durante la etapa embrionaria se activan centros de señalización: uno similar al «cerebro anterior» (prosencéfalo), en la trompa (proboscide); uno similar al «cerebro medio» (mesencéfalo) en el collar de tejido en la unión de la trompa y el collar y uno similar al «cerebro posterior» (rombencéfalo) que se encuentra donde el collar se une al tronco del gusano S. kowalevskii.[10][11]
Las proteínas de señalización en cada centro interactúan unas con otras en forma muy parecida a como lo hacen en los vertebrados. Expresan como los vertebrados las tres combinaciones de proteínas (ANR, ZLI e IsO), en regiones comparables del embrión durante el desarrollo de sus tres partes del cerebro.
Comparten una arquitectura de regulación de transcripción ampliamente conservada durante el patrón corporal temprano.[12][9][13]
Véase también[editar]
Referencias[editar]
- ↑ «Saccoglossus kowalevskii». WoRMS (en inglés). Consultado el 25 de octubre de 2023.
- ↑ a b Gonzalez P.; Uhlinger K.R.; Lowe C.J. (2016). «The Adult Body Plan of Indirect Developing Hemichordates Develops by Adding a Hox-Patterned Trunk to an Anterior Larval Territory». Current Biology (en inglés) 27 (1): 87-95. doi:10.1016/j.cub.2016.10.047. Consultado el 25 de octubre de 2023.
- ↑ a b Kaul-Strehlow S.; Stach T. (2013). «A detailed description of the development of the hemichordate Saccoglossus kowalevskii using SEM, TEM, Histology and 3D-reconstructions». Frontiers in Zoology (en inglés) 10 (53). Consultado el 25 de octubre de 2023.

- ↑ Miller D.C. (1992). «Response of Saccoglossus kowalevskii (Phylum Hemichordata, Class Enteropneusta) to changes in diet». Marine ecology progress series (en inglés) 87: 41-54.
- ↑ Simakov O. (2015). «Hemichordate genomes and deuterostome origins» (en inglés). Consultado el 25 de octubre de 2023.
- ↑ a b Robinson R. (2006). «Unique Development in Hemichordates Suggests Some Unique Features of Chordates». PLoS Biology (en inglés) (Public Library of Science) 4 (9): e288. doi:10.1371/journal.pbio.0040288. Consultado el 2 de noviembre de 2023.
- ↑ Lowe C.; Terasaki M.; Wu M.; Freeman R.M. Jr.; Runft L.; Kwan K.; Haigo S.; Aronowicz J.; Lander E.; Gruber C.; Smith M.; Kirschner M.; Gerhart J. (2006). «Dorsoventral Patterning in Hemichordates: Insights into Early Chordate Evolution.». PLoS Biology (en inglés) (Public Library of Science) 4 (9): e291. doi:10.1371/journal.pbio.0040291. Consultado el 31 de octubre de 2023.
- ↑ a b Kerbl A.; Jékely G. (2023). «Hemichordates’ diffuse “skin brain” shows unexpected complexity». PLoS Biology (en inglés) (Public Library of Science) 21 (9): e3002312. doi:10.1371/journal.pbio.3002312. Consultado el 8 de noviembre de 2023.
- ↑ a b Andrade López J.M.; Pani A.M.; Wu M.; Gerhart J.; Lowe C.J. (2023). «Molecular characterization of nervous system organization in the hemichordate acorn worm Saccoglossus kowalevskii.». PLoS Biology (en inglés) (Public Library of Science) 21 (9): e3002242. doi:10.1371/journal.pbio.3002242. Consultado el 25 de octubre de 2023.
- ↑ «El origen secreto del cerebro de los vertebrados». Vista al Mar. 2012.
- ↑ Maxmen, Amy (2012). «Marine worm rewrites theory of brain evolution». Nature News (en inglés). Consultado el 25 de octubre de 2023.

- ↑ Pani A.M.; Mullarkey E.E.; Aronowicz J.; Assimacopoulos S.; Grove E.A.; Lowe C.J. (2012). «Ancient deuterostome origins of vertebrate brain signalling centres». Nature (en inglés) 483: 289-294. Consultado el 25 de octubre de 2023. (requiere suscripción).
- ↑ «El genoma del gusano de bellota revela los orígenes branquiales de la faringe humana». Science. 2015. Consultado el 25 de octubre de 2023.
Enlaces externos[editar]
- El origen secreto del cerebro de los vertebrados. Vista al Mar, (2012).
- Los orígenes branquiales de la faringe humana Science, (2015).